 "
"
Team:British Columbia/modeling description
From 2010.igem.org
Introduction
We developed a mathematical model that describes the dynamics of the biofilm structure (in terms of bacterial population size) and the interactions among the major components, including the engineered phage and DispersinB (DspB) protein. We used numerical simulations to predict the impact of phage and DspB release on the biofilm structure. We also investigated the weight of each parameter to the design of our system with sensitivity analysis. Possible scenarios of biofilm degradation suggested by our model are explored.
Model Description
Basic Biofilm Geometry
The biofilm system assumes a simple planar geometry characterized by depth, D, and cross-sectional area, A (Figure 1). The density and distribution of the biofilm bacterial population, extracellular polymeric substance (EPS), and dissolved components (e.g. AIP and metabolites) are uniform throughout the biofilm structure. Assuming that each biofilm bacterium occupies a spherical volume of diameter, d, and the surrounding EPS extends this volume by a constant, α, each bacterium takes up a cubic volume of (α + d)3. The total biofilm population in a biofilm can be estimated:
Biofilm Bacteria
The total bacterial population, B, is divided into two subpopulations: 1) the carrier bacteria, Bi, which are infected with the engineered phage and 2) the non-carrier bacteria, Bu, which are uninfected but susceptible to phage infection upon exposure. We separate the carrier bacteria subpopulation further into two subpopulations: 1) the bacteria infected with the phage in latent phase, Bl, and 2) those infected with the phage in lytic phase, BL. The total population, BT, undergo logistic growth at rate ρT limited by the carrying capacity, K; the uninfected, Bu, and infected, Bi, subpopulations grow at rates ρu and ρi, respectively. The following differential equations describe the dynamics of the two infected subpopulations, where p is the portion of latent bacteria, π the rate of transition from latent to lytic phase, and λ the rate of lysis:
EquationsThe carrying capacity determines the maximum biofilm thickness. The carrying capacity can be coarsely estimated using Equation 1 by letting D equal to the maximum thickness.
Phage Particles
Initially, only the engineered S. aureus bacteria will be introduced to the biofilm. In response to the presence of AIP, they will generate and release the first batch of phage particles. A proportion of these phage particles, κ, will successfully infect and integrate its genetic information into the host genome. Once infected, a bacterium is subject to lysis probability of λ; upon lysis, the bacterium will release R number of phage particles. The dynamics of the phage particles is governed by half-life, t1/2.
Phage Diffusion and Invasion
Lysis of the infected host cells forms a pool of newly produced phage particles. This phage population diffuses out 1) towards the bulk liquid at rate rout or 2) into the biofilm structure through EPS at rate rin. We assume that the new phage pool is concentrated in a defined layer immediately after lysis. This layer serves as the initial point of diffusion. The diffusion of phage particles into the biofilm can be modeled by Fick’s second law of diffusion, where φ is the phage concentration and x the distance from the concentrated layer:
EquationsLet us redefine the concentration, φ, to be relative to the initial phage concentration, φ0, at time t = 0 such that EQUATION: Equations
Solving for φr, we derive:
The distance of phage diffusion within one time step (i.e. from t = 0 to t = 1) can be estimated by letting φr = 0 and solving for x, which is dependent on the constant, θ, and diffusion coefficient, rin (Equation 10.1) Note that the integral cannot be solved analytically in closed form. Numerical methods such as the adaptive Simpson quadrature are required to estimate it.
EquationsSince we are treating this as a one-dimensional problem, the distance of phage diffusion can be roughly estimated by the diffusion length for a certain time period Δt:
For simplicity, we will implement this equation in our simulations.
EquationsThis distance reflects the depth of the phage-infested layer and is related to the proportion of the total biofilm susceptible to infection. We simplify this relationship by using information about the geometry of the biofilm bacteria and biofilm structure. The following represents the relationship between the depth of the remaining biofilm (i.e. not disintegrated by phage) and the total biofilm population.
DspB Activity
The production of DspB enzymes is coupled with phage production since they are both activated via the same signal pathway. The concentration of DspB enzymes in the EPS is subject to diffusion forces similar to those acting on the phage particles. The dynamics of extracellular DspB concentration is therefore, where S is the amount of DspB released per host lysis:
EquationThe lysis release of DspB enzymes promotes phage invasion by facilitating access into the biofilm structure due to their EPS-degrading activity. Therefore, DspB increases the diffusion rate of the phage particles into biofilm. The diffusion rate is related to the concentration of DspB in the phage-infested layer, where Y is the proportionality constant:
AIP Activity
Biofilms maintain their extracellular concentration of AIP at . Here, we assume that AIP is being secreted by the biofilm bacteria at rate which is a function of the total biofilm population and degrades at rate γ. The dynamics of AIP concentration is assumed to behave more or less logistically:
EquationThe activation of phage production and dspB transcription is dependent on the extracellular concentration of AIP. We engineered the system so that it responds to a certain threshold level of AIP, [AIP]thres, below which phage production and dspB transcription ceases (i.e. ). Modifying Equation 12, we derive conditional differential equations describing phage dynamics:
EquationFurthermore, AIP-dependent dynamics of DspB production entails, where when extracellular AIP concentration falls below the activation threshold. Since dspB transcription is coupled with phage generation, we derive differential equations of DspB dynamics similar to those of phage dynamics:
Output Summary
The progress and outcome of phage invasion can be monitored by the populations of the biofilm and the phage. Hence, the output of the model can be summarized by using various measures incorporating the variables BT, and P:
1) percent of biofilm population remaining since phage introduction (Bp), where B0 is the initial biofilm population:
2) viral infection potential (VIP):
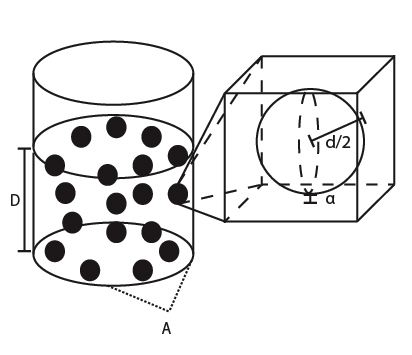
Figure 1: Schematic diagram of the biofilm structure. A cylindrical (or rectangular) container of depth, D, and cross-sectional area, A, holds the biofilm mass. Each bacterium occupies a spherical volume of diameter, d, corrected by a constant representing the space filled by surrounding EPS, α (see zoom-in).

Figure 2: Flow diagram indicating the interactions among the components of the biofilm system. The total biofilm population, BT, is divided into subpopulations (inside the rounded box). The phage particle population, P, directly interacts with only the subpopulation infected with latent phage, BL.

Figure 3: Schematic diagram of the phage invasion process. Phage particles move into the biofilm structure via simple diffusion. Lysis of infected host cells in the phage-infested layer (indicated by a thickness of x) produces new pools of phage particles which undergo diffusion into the biofilm at rate, rin, or out of biofilm at rate ,rout. The depth of the biofilm structure is z.