"
"
Team:MIT mammalian Switch
From 2010.igem.org
| (14 intermediate revisions not shown) | |||
| Line 25: | Line 25: | ||
<ul> | <ul> | ||
<li><a href="https://2010.igem.org/Team:MIT_toggle">Overview</a></li> | <li><a href="https://2010.igem.org/Team:MIT_toggle">Overview</a></li> | ||
| + | <li><a href="https://2010.igem.org/Team:MIT_tmodel">Modelling</a></li> | ||
<li><a href="https://2010.igem.org/Team:MIT_tconst">Toggle Construction</a></li> | <li><a href="https://2010.igem.org/Team:MIT_tconst">Toggle Construction</a></li> | ||
| - | <li><a href=" | + | <li><a href="https://2010.igem.org/Team:MIT_composite">Characterization</a></li> |
</ul> | </ul> | ||
</dd> | </dd> | ||
| Line 60: | Line 61: | ||
<div id="unique" style="padding:0px; font-size: 14px; border: 1px solid black; margin:0px; background-color:transparent;"> | <div id="unique" style="padding:0px; font-size: 14px; border: 1px solid black; margin:0px; background-color:transparent;"> | ||
| - | <table width=650px style="background-color: white; margin-top:5px; padding: 10px;" | + | <table width=650px style="background-color: white; margin-top:5px; padding: 10px;"> |
| - | <tr><td> | + | <tr><td> |
| + | <div class="outline"> | ||
| + | <a href="#why">1 Why Synthetic Switches?</a><br> | ||
| + | <a href="#bistable">2 How Positive Feedback Leads to Bistability</a><br> | ||
| + | <a href="#simp">3 Our Simplified Toggle</a><br> | ||
| + | <ul><a href="#results"> Results </a><br></ul> | ||
| + | <a href="#full">4 Our Full Toggle</a><br> | ||
| + | <ul><a href="#model"> Model </a><br></ul> | ||
| + | </div></td><tr><td> | ||
| - | < | + | <div class="bodybaby">Synthetic Switch: Bistable Toggles in Mammalian Cells</div></td></tr> |
| + | |||
| + | <tr><td><b class="bolded" id="why">WHY SYNTHETIC SWITCHES?</b> | ||
| + | |||
| + | <p>It is useful for a synthetic system capable of sensing mechanical stimuli to be able to convert a transient signal into a constitutive response. We want our system to sense mechanical stimuli for a period of time, during which an internal switch is flipped from the "off" state to the "on" state. After the removal of the stimulus, the cell will stay in the "on" state, during which it secretes BMP2 and osteogenesis occurs. Because cell differentiation occurs on a much longer time scale than desired mechanical stimulation, such a switch is required for a system to be of use. | ||
| + | <HR> | ||
| + | <p><b class="bolded" id="bistable">HOW POSITIVE FEEDBACK CAN BE USED TO CONSTRUCT A BISTABLE TOGGLE</b> | ||
<p> | <p> | ||
| + | <a href="https://static.igem.org/mediawiki/2010/b/b6/Bistable-Toggles.png" class="thickbox" title="How positive Feedback-->Bistable Toggle. In our system, protein A is analogous to rtTA3/rtTA, and promoter A is analogous to TREt."><img src="https://static.igem.org/mediawiki/2010/b/b6/Bistable-Toggles.png" width=300px></a> | ||
| + | <p>Consider a transcriptional positive feedback system, in which an outside signal causes the transcription of an inducer of transcription, protein A, which induces the transcription and production of more protein A. When the signal is added, transcription and translation of protein A increases, which causes its own transcription to increase further. The outside stimulating signal reaches a threshold value such that the removal of the signal does not affect the level of protein A production, because at this point almost all the transcription of protein A is driven by protein A, and not the outside signal. At this point we say the cell has reached a new high output state. This state is stable down to a different threshold value of protein A than the previous threshold. Thus, using positive feedback loops, we can construct a bistable toggle with two stable states: low transcription, and high transcription. | ||
| + | |||
| + | <HR> | ||
| + | <p><b class="bolded" id="simp">SIMPLIFIED BISTABLE TOGGLE</b> | ||
| + | <p>During the summer we designed both a complete and a simplified bistable toggle with a low and high output state using positive feedback loops. | ||
| + | <a href=https://static.igem.org/mediawiki/2010/1/1d/Simplified.png><img src="https://static.igem.org/mediawiki/2010/1/1d/Simplified.png" width=100%></a> | ||
| + | <p><ul> | ||
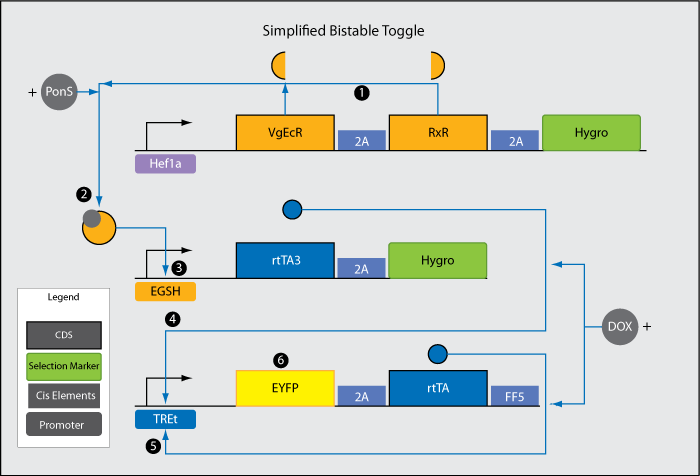
<br>1. Hef1a promoter leads to low level, constitutive expression of the two halves (VgEcR and RxR) of a receptor responsive to the chemical ponasterone A (PonS). | <br>1. Hef1a promoter leads to low level, constitutive expression of the two halves (VgEcR and RxR) of a receptor responsive to the chemical ponasterone A (PonS). | ||
<br>2. VgEcR, RxR, and PonS combine to form an active complex. | <br>2. VgEcR, RxR, and PonS combine to form an active complex. | ||
| Line 75: | Line 98: | ||
<li>Note 1: DOX in this system would be already present in the media. rtTA3 combines with DOX to form an activated complex in order to induce TREt. The outside signal being added is PonS. Without PonS addition, the presence of DOX is unable to flip the switch because there is very low level transcription of rtTA3. | <li>Note 1: DOX in this system would be already present in the media. rtTA3 combines with DOX to form an activated complex in order to induce TREt. The outside signal being added is PonS. Without PonS addition, the presence of DOX is unable to flip the switch because there is very low level transcription of rtTA3. | ||
<li>Note 2: 2A sites are cis elements that enable polycistronic translation in mammalian cells. Proteins before and after the 2A site are translated at roughly equal frequency. | <li>Note 2: 2A sites are cis elements that enable polycistronic translation in mammalian cells. Proteins before and after the 2A site are translated at roughly equal frequency. | ||
| - | <br> | + | <br> </ul> |
| - | </ | + | <HR> |
| - | < | + | <p><b class="bolded" id="results">Results</b> |
| - | + | ||
| - | </ | + | |
| + | <p><img src=https://static.igem.org/mediawiki/2010/2/2c/Sensitivity_analysis.gif> | ||
| + | <br> Figure 1. Effect of DOX Concentration on Bistability of System. Time lapse shows effect of DOX on the system, where 3 x intercepts are required for bistability. Sensitivity analysis of rate plot shows that DOX concentration is critical to achieving a bistable system. | ||
| - | < | + | <p><img src=https://static.igem.org/mediawiki/2010/a/a8/Bar-graph-01-and-1x-DOX.png width=300> |
| + | <br> Figure 2. Fluorescence output from the toggle was measured post transient transfection of HEK293FT cells with the toggle construct, in response to PonS. High DOX concentration show no sensitivity to PonS. Low DOX concentration show sensitivity to PonS. | ||
| + | <p>The figure below show a preliminary experiment during which PonS was added post transient calcium phosphate transfection of a HEK293FT cell line containing the Hef1a_VgEcR_RxR construct with the other two components of the toggle. | ||
| + | </tr> | ||
| + | <tr><td><img src=https://static.igem.org/mediawiki/2010/d/dc/Toggle-fl-figure.png> | ||
| + | <br>Figure 3. Induction of EYFP Expression in High Output State in Mammlian Toggle. | ||
| + | <ul><p>There is significant increase in fluorescence in the cells transfected with the toggle with addition of PonS in the system. Red fluorescence indicates transfection efficiency. EYFP indicates output from the toggle. | ||
| + | |||
| + | </td> | ||
| + | <tr><td><HR> | ||
| + | <b class="bolded" id="full">FULL MAMMALIAN TOGGLE</b> | ||
| + | <p>We also designed a full bistable toggle designed to drastically shorten the off-->on and on-->off switch via microRNA. Arvind, a member of the iGEM team, has mathematically modeled this circuit in depth, along with several others that were also considered. | ||
| + | <HR> | ||
| + | <b class="bolded" id="model"> | ||
| + | <br>Please view the model <a href=https://static.igem.org/mediawiki/2010/e/e0/Mammalian_Full_Toggle.pdf>here</a>.</b> | ||
</td></tr></table> | </td></tr></table> | ||
Latest revision as of 21:18, 17 November 2010
|
Synthetic Switch: Bistable Toggles in Mammalian Cells |
| WHY SYNTHETIC SWITCHES?
It is useful for a synthetic system capable of sensing mechanical stimuli to be able to convert a transient signal into a constitutive response. We want our system to sense mechanical stimuli for a period of time, during which an internal switch is flipped from the "off" state to the "on" state. After the removal of the stimulus, the cell will stay in the "on" state, during which it secretes BMP2 and osteogenesis occurs. Because cell differentiation occurs on a much longer time scale than desired mechanical stimulation, such a switch is required for a system to be of use. HOW POSITIVE FEEDBACK CAN BE USED TO CONSTRUCT A BISTABLE TOGGLE
Consider a transcriptional positive feedback system, in which an outside signal causes the transcription of an inducer of transcription, protein A, which induces the transcription and production of more protein A. When the signal is added, transcription and translation of protein A increases, which causes its own transcription to increase further. The outside stimulating signal reaches a threshold value such that the removal of the signal does not affect the level of protein A production, because at this point almost all the transcription of protein A is driven by protein A, and not the outside signal. At this point we say the cell has reached a new high output state. This state is stable down to a different threshold value of protein A than the previous threshold. Thus, using positive feedback loops, we can construct a bistable toggle with two stable states: low transcription, and high transcription. SIMPLIFIED BISTABLE TOGGLE During the summer we designed both a complete and a simplified bistable toggle with a low and high output state using positive feedback loops.
1. Hef1a promoter leads to low level, constitutive expression of the two halves (VgEcR and RxR) of a receptor responsive to the chemical ponasterone A (PonS). 2. VgEcR, RxR, and PonS combine to form an active complex. 3. EGSH is induced by the VgECR-RxR-PonS complex. 4. rtTA3 is transcribed after teh activation of EGSH. 5. Each rtTA3 binds to two of the six tetO sites in the TREt promoter and induces the promoter. 6. rtTA, another version of rtTA3, further induces activation of the TREt promoter. This establishes a positive feedback loop, which propels the system from a low to a stable high output state (the switch is "flipped"). In the high output state, EYFP is constitutively expressed at a high level. Results
The figure below show a preliminary experiment during which PonS was added post transient calcium phosphate transfection of a HEK293FT cell line containing the Hef1a_VgEcR_RxR construct with the other two components of the toggle. |

Figure 3. Induction of EYFP Expression in High Output State in Mammlian Toggle. There is significant increase in fluorescence in the cells transfected with the toggle with addition of PonS in the system. Red fluorescence indicates transfection efficiency. EYFP indicates output from the toggle. |
FULL MAMMALIAN TOGGLE We also designed a full bistable toggle designed to drastically shorten the off-->on and on-->off switch via microRNA. Arvind, a member of the iGEM team, has mathematically modeled this circuit in depth, along with several others that were also considered. Please view the model here. |