 "
"
Team:INSA-Lyon/Project/Stage3/Theory
From 2010.igem.org
m |
|||
| (61 intermediate revisions not shown) | |||
| Line 5: | Line 5: | ||
{{INSA-Lyon/menugauche}} | {{INSA-Lyon/menugauche}} | ||
| + | {{INSA-Lyon/menuImage}} | ||
| + | {{INSA-Lyon/menudroppy3}} | ||
<html xmlns="http://www.w3.org/1999/xhtml" dir="ltr" lang="en-US" xml:lang="en"> | <html xmlns="http://www.w3.org/1999/xhtml" dir="ltr" lang="en-US" xml:lang="en"> | ||
| Line 14: | Line 16: | ||
} | } | ||
ul#topnav li span { background: #f200d0;} | ul#topnav li span { background: #f200d0;} | ||
| - | .image {width: | + | .image {width:350px; height:263px; margin-top: 10px; margin-bottom: 5px; position: relative;float : left} |
| - | .image1 {width: | + | .imagea {width:370px; height:263px; margin-top: 10px; margin-bottom: 5px; position: relative;float : right} |
| + | .image1 {width:200px; height:140px; margin-top: 10px; margin-bottom: 5px; position: relative;} | ||
.image2 {width:722px; height:147px; margin-top: 10px; margin-bottom: 5px; position: relative;} | .image2 {width:722px; height:147px; margin-top: 10px; margin-bottom: 5px; position: relative;} | ||
.image3 {width:596px; height:112px; margin-top: 10px; margin-bottom: 5px; position: relative;} | .image3 {width:596px; height:112px; margin-top: 10px; margin-bottom: 5px; position: relative;} | ||
| - | |||
| + | #list_extra{list-style-type:circle; margin: 10px auto auto 100px;} | ||
| + | </style> | ||
<script type="text/javascript"> | <script type="text/javascript"> | ||
$(document).ready(function() { | $(document).ready(function() { | ||
| Line 34: | Line 38: | ||
</script> | </script> | ||
| - | + | <a href="top"></a> | |
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | <div id=" | + | <div id="corps2"> |
| - | < | + | <h2>Regulation</h2> |
| - | < | + | <br> |
| - | < | + | <p>The system of protein purification needs to be regulated to be completely operational. In fact, the cell machinery has limited energy sources to cope with the DNA replication, proliferation, and simultaneously, the granules and proteins synthesis. We need to space out the synthesis of the different other stages.</p> |
| - | </ | + | <br /> |
| + | |||
| + | <p>As described by Banki et al.(Protein Science, 2005), proteins can bind directly to the granules already shaped. Moreover the protein synthesis rate will be higher if the granules synthesis is stopped. Thus we planned to induce the granule synthesis during 30 hours before turning it off and starting the protein synthesis. So we needed promoters which turn each other ON and OFF. We designed two distinct systems in order to have an alternative if one of them didn't work as expected. | ||
| + | |||
| + | </p> | ||
| + | <br /><br /> | ||
| + | <h3><font color="purple">Thermoregulation</font></h3><br> | ||
| + | |||
| + | |||
| + | <br /> | ||
| + | <p>First, we wanted to take advantage of our curli promoter which has the ability to be switched ON at 28°C and OFF at higher temperatures. You can read <a href="https://2010.igem.org/Team:INSA-Lyon/Project/Stage3/Strategy/Theorycurli"> the theory </a> concerning curli and ompR to understand how this promoter and its regulators work.<br/></p> | ||
| + | <p> | ||
| + | Then, we looked for an iGEM promoter also regulated by temperature, we chose a thermometer RNA. | ||
| + | It is switched ON above 32°C, allowing the transcription (<a href="http://partsregistry.org/Part:BBa_K115017"> BBa_K115017</a>). | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p>When both functional constructs are in the same cell, the culture is moved to 28°C so the PHB granules are synthesized under the control of Curli promoter. Then, the culture is moved to 37°C allowing the synthesis of the target proteins, activated by the thermometer RNA. At 37°C, the Curli promoter is turned OFF and the PHB granules are not synthesized anymore.</p> | ||
| + | |||
| + | |||
| + | <div style="text-align:left;"> | ||
| + | <div style="text-align:right;"> | ||
| + | <div style="text-align:left;"> | ||
| + | |||
| + | |||
| + | <SCRIPT LANGUAGE="JavaScript"> | ||
| + | <!-- | ||
| + | imageNormal=new Image | ||
| + | imageNormal.src="http://lh6.ggpht.com/_Uc3bmii-yi0/TMh7jvSH4UI/AAAAAAAAApA/3AMef32byTc/thRegu28C.PNG" | ||
| + | image28=new Image | ||
| + | image28.src="http://lh6.ggpht.com/_Uc3bmii-yi0/TMh7jvSH4UI/AAAAAAAAApA/3AMef32byTc/thRegu28C.PNG" | ||
| + | image37=new Image | ||
| + | image37.src="http://lh3.ggpht.com/_Uc3bmii-yi0/TMh7jwizLUI/AAAAAAAAApE/tFX1wz4Tp7s/thRegu37C.PNG" | ||
| + | //--> | ||
| + | </SCRIPT> | ||
| + | |||
| + | |||
| + | <div class="colonnegauche"> | ||
| + | <div id="menuImage"> | ||
| + | <ul> | ||
| + | <li name="28" | ||
| + | ONMOUSEOVER="document.image1.src=image28.src" | ||
| + | ONMOUSEOUT="document.image1.src=imageNormal.src"> | ||
| + | <a class="brn"> > At 28°C</a></li> | ||
| - | <li | + | <li name="37" |
| - | + | ONMOUSEOVER="document.image1.src=image37.src" | |
| - | + | ONMOUSEOUT="document.image1.src=imageNormal.src"> | |
| - | + | <a class="orange"> > At 37°C</a></li> | |
| - | + | </ul> | |
| - | + | ||
</div> | </div> | ||
</div> | </div> | ||
| + | <IMG id="image" NAME="image1" SRC="http://lh6.ggpht.com/_Uc3bmii-yi0/TMh7jvSH4UI/AAAAAAAAApA/3AMef32byTc/thRegu28C.PNG" alt="Theory Regulation" style="margin-left:20px; margin-right:30px;"/> | ||
| + | <IMG id="image" NAME="imagelegend" SRC="http://lh6.ggpht.com/_Uc3bmii-yi0/TMh7jxvKr0I/AAAAAAAAApI/8wc5Y0fiVhU/thReguLegend.PNG" alt="Theory Regulation Legend"/> | ||
| - | <div | + | </div> |
| - | < | + | |
| - | + | <p style="font-size:0.9em; text-indent:0px; text-align:center;"><em></em></p> | |
| - | + | ||
| - | + | ||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
<br /> | <br /> | ||
<br /> | <br /> | ||
<br /> | <br /> | ||
| + | <br/> | ||
| + | <h3><font color="purple">Control under Arabinose</font></h3><br> | ||
| - | |||
| - | |||
| - | <p> | + | <br> |
| + | <p>In an other way, we wanted to use the combination of the promoter pBad/araC and the promoter LuxR/cI.</p> <br> | ||
| + | |||
| + | <p>Without arabinose, LuxR and LuxI proteins are synthesized constitutively. LuxI is involved in the synthesis of HomoSerine Lactone (HSL). The fixation of HSL in LuxR protein causes a conformational change of the protein. In this conformation, LuxR protein can interact with the promoter and activates it. This leads to PHB synthesis and when PHB molecules are accumulated enough, they organize themselves in granules. | ||
| + | The promoter Pbad/araC, without arabinose, is in a off state. | ||
</p> | </p> | ||
| + | <p>When arabinose is added to the medium, it interacts with Pbad/araC and induces the synthesis of cI protein and phasin-intein construction. cI protein negatively regulates the promoter LuxR/HSL and stops PHB synthesis. Its effect is more efficient than the effect of the LuxR/HSL complex. Therefore, the promoter regulated by LuxR/HSL is totally turned off. During this step, phasin and intein proteins are synthesized and get into PHB granules.<br><br> </p><br> | ||
| + | |||
<div style="text-align:center;"> | <div style="text-align:center;"> | ||
| - | <img | + | <img src="http://lh3.ggpht.com/_Uc3bmii-yi0/TMh-oPnKpUI/AAAAAAAAApc/2DNV5QBTPPs/avecetsansArabinose.PNG" alt="With and without Arabinose" title="With and without Arabinose"/> |
| - | alt=" | + | |
</div> | </div> | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
<div style="text-align:center;"> | <div style="text-align:center;"> | ||
| - | <img | + | <img src="http://lh6.ggpht.com/_Uc3bmii-yi0/TMdHOjqQmRI/AAAAAAAAAjg/lQaP7d5dFok/ArabinoseLegend.PNG" /> |
| + | </div> | ||
| + | <br> | ||
| + | |||
| + | </div> | ||
</div> | </div> | ||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
</div> | </div> | ||
| - | |||
| - | + | <p style="text-align:center;"><a href="#top">Top of Page</a></p> | |
| - | </ | + | |
<div id="corps4" style="border:0px;"> | <div id="corps4" style="border:0px;"> | ||
Latest revision as of 19:34, 27 October 2010
Regulation
The system of protein purification needs to be regulated to be completely operational. In fact, the cell machinery has limited energy sources to cope with the DNA replication, proliferation, and simultaneously, the granules and proteins synthesis. We need to space out the synthesis of the different other stages.
As described by Banki et al.(Protein Science, 2005), proteins can bind directly to the granules already shaped. Moreover the protein synthesis rate will be higher if the granules synthesis is stopped. Thus we planned to induce the granule synthesis during 30 hours before turning it off and starting the protein synthesis. So we needed promoters which turn each other ON and OFF. We designed two distinct systems in order to have an alternative if one of them didn't work as expected.
Thermoregulation
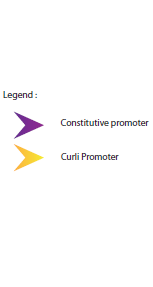
First, we wanted to take advantage of our curli promoter which has the ability to be switched ON at 28°C and OFF at higher temperatures. You can read the theory concerning curli and ompR to understand how this promoter and its regulators work.
Then, we looked for an iGEM promoter also regulated by temperature, we chose a thermometer RNA. It is switched ON above 32°C, allowing the transcription ( BBa_K115017).
When both functional constructs are in the same cell, the culture is moved to 28°C so the PHB granules are synthesized under the control of Curli promoter. Then, the culture is moved to 37°C allowing the synthesis of the target proteins, activated by the thermometer RNA. At 37°C, the Curli promoter is turned OFF and the PHB granules are not synthesized anymore.

Control under Arabinose
In an other way, we wanted to use the combination of the promoter pBad/araC and the promoter LuxR/cI.
Without arabinose, LuxR and LuxI proteins are synthesized constitutively. LuxI is involved in the synthesis of HomoSerine Lactone (HSL). The fixation of HSL in LuxR protein causes a conformational change of the protein. In this conformation, LuxR protein can interact with the promoter and activates it. This leads to PHB synthesis and when PHB molecules are accumulated enough, they organize themselves in granules. The promoter Pbad/araC, without arabinose, is in a off state.
When arabinose is added to the medium, it interacts with Pbad/araC and induces the synthesis of cI protein and phasin-intein construction. cI protein negatively regulates the promoter LuxR/HSL and stops PHB synthesis. Its effect is more efficient than the effect of the LuxR/HSL complex. Therefore, the promoter regulated by LuxR/HSL is totally turned off. During this step, phasin and intein proteins are synthesized and get into PHB granules.

