 "
"
Team:Queens-Canada/nervous
From 2010.igem.org
| Line 167: | Line 167: | ||
<html><a name="gpcr_table"></a><div class="section"><h2>G-Protein–Coupled Receptors (GPCRs)</h2></html> | <html><a name="gpcr_table"></a><div class="section"><h2>G-Protein–Coupled Receptors (GPCRs)</h2></html> | ||
| + | |||
| + | <html><center><img src="http://upload.wikimedia.org/wikipedia/commons/thumb/c/c9/GPCR_cycle.jpg/800px-GPCR_cycle.jpg" style="width: 100%; max-width: 800px; box-shadow: 1px 2px 3px #808080"><br> | ||
| + | <a target="_new" href="http://en.wikipedia.org/wiki/G_protein-coupled_receptor">From <i>Wikipedia</i></a></center></html> | ||
GPCRs are common features of eukaryotes and, in multicellular organisms, are one of the major means by which tissues communicate with one another. In ''C. elegans'', they are also used for sensory purposes via the amphid. (We use some of them similarly in our noses.) Many projects have the potential to be highly dependent on GPCR importation and engineered GPCRs, although so far their use at iGEM has not been extensive owing to the high amount of work required to get them to function in prokaryotes. | GPCRs are common features of eukaryotes and, in multicellular organisms, are one of the major means by which tissues communicate with one another. In ''C. elegans'', they are also used for sensory purposes via the amphid. (We use some of them similarly in our noses.) Many projects have the potential to be highly dependent on GPCR importation and engineered GPCRs, although so far their use at iGEM has not been extensive owing to the high amount of work required to get them to function in prokaryotes. | ||
Latest revision as of 21:54, 27 October 2010
The Nervous System
The Amphid
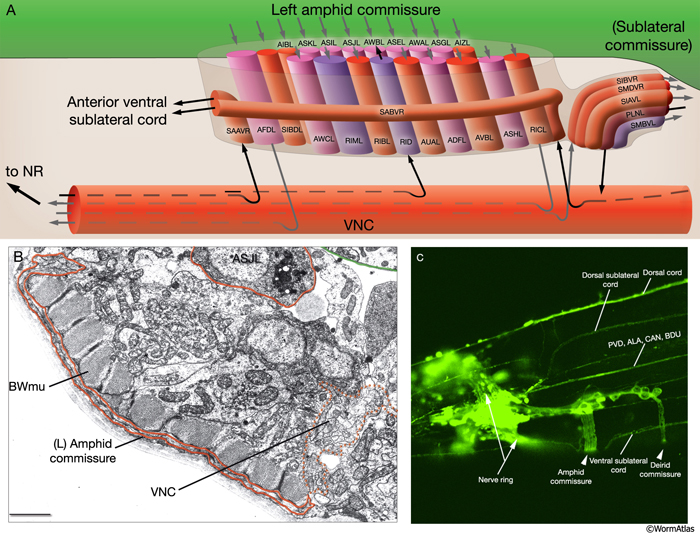
From WormAtlas
The amphid of C. elegans is equivalent (and homologous) to the nose of a human. It is used for sensing volatile and water soluble chemicals, and is found in the anterior region of the worm, at the base of the lips. Consisting of 12 sensory neurons, some of which are ciliated, the amphid is the primary sensory organ of C. elegans.
There are only two responses that C. elegans has to olfactory stimuli: attraction and repulsion. These are governed by separate sensory neurons, which are hard-wired upstream to produce those responses, and use largely interchangeable sensors in the form of receptor proteins. The majority of these receptors are GPCRs and membrane-bound guanylyl cyclases. It has been shown that adding new GPCRs can create either reaction depending on where the GPCR is expressed, and that at least some GPCRs from humans and mice will work in C. elegans without modification.
There are a large number of GPCRs and receptor GCs that occur natively in C. elegans; over one thousand GPCR candidates have been identified through genomic analysis. However, their functions and stimuli are not well understood.
The primary appeal of engineering the amphid is to add new receptors so that the worm is either attracted to, or repulsed by, some sensory input. The worm has very powerful chemotaxis, and could effectively replace bacteria in most biosensor applications. Below is a partial table from WormBook listing the neurons of the amphid, their functions, and some of the receptors that occur in each. The gpa family encodes not receptors, but G-protein α subunits, which are required for signal transduction from GPCRs and are the major influential factor in whether or not a specific GPCR can be used in a given neuron without modification.
| Neuron | Function | Receptors; G proteins |
| ASE | Water-soluble chemotaxis | Receptor guanylate cyclases?; gpa-3 |
| AWC | Volatile chemotaxis, Lifespan, Navigation | GPCRs (str-2); odr-3 (major), gpa-3, gpa-2, gpa-5, gpa-13 |
| AWA | Volatile chemotaxis, Lifespan (minor) | GPCRs (odr-10); odr-3 (major), gpa-3, gpa-5; gpa-13; gpa-6 |
| AWB | Volatile avoidance | GPCRs; odr-3 |
| ASH | Nociception: Osmotic avoidance, Nose touch avoidance, Chemical avoidance, Social feeding | GPCRs; odr-3 (major), gpa-3 (major), gpa-11, gpa-1, gpa-13, gpa-14, gpa-15 |
| ASI | Dauer formation, Chemotaxis(minor), Navigation | GPCRs; gpa-1, gpa-3, gpa-4, gpa-5, gpa-6, gpa-10, gpa-14 |
| ADF | Dauer formation, Chemotaxis (minor) | GPCRs; odr-3, gpa-3, gpa-10, gpa-13 |
| ASG | Dauer formation (minor), Lifespan, Chemotaxis (minor) | GPCRs; gpa-3 |
| ASJ | Dauer formation and recovery, Chemotaxis (minor), Lifespan | GPCRs; gpa-1, gpa-3, gpa-9, gpa-10, gpa-14 |
| ASK | Avoidance (minor), Chemotaxis (minor), Lifespan, Navigation | GPCRs; gpa-2, gpa-3, gpa-14, gpa-15 |
| ADL | Avoidance (minor), Social feeding | GPCRs; gpa-1, gpa-3, gpa-11, gpa-15 |
| URX, AQR, PQR | Oxygen/aerotaxis, Social feeding | Soluble guanylate cyclases (gcy-35, gcy-36); gpa-8 |
| PHA, PHB | Avoidance (antagonistic) | GPCRs; gpa-1, gpa-2, gpa-3, gpa-6, gpa-9, gpa-13, gpa-14, gpa-15 |
The major object of desire when working with sensory neurons is the availability of a specific promoter that can target each individually. We have endeavored to provide a number of these; see our parts list. More detail on chemosensation in C. elegans is available from WormBook.
Other Forms of Sensation
C. elegans is also natively able to detect mechanical force exerted on its body as well as heat and cold. The potential applications for engineering these traits is minimal, although it is worth noting that C. elegans naturally moves towards temperatures at which it has known feeding conditions to be favorable, and away from those at which it was not able to feed. There are, however, mutants that habitually seek out cold (cryophilic) and heat (thermophilic) without regard for this process. These may be of use, and can be found in our article on getting strains.
The Neuron
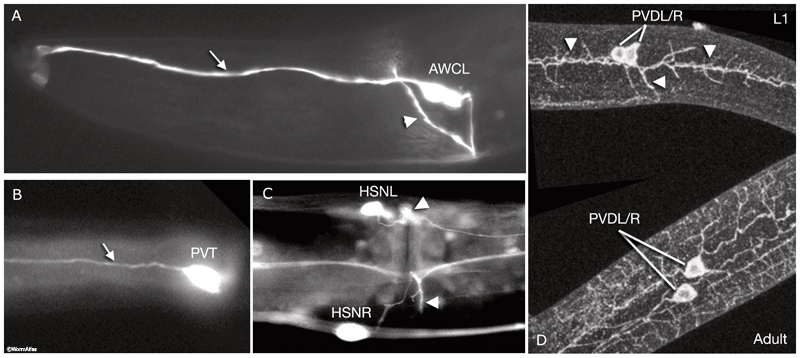
From WormAtlas
The neuron is a highly-specialized cell that provides the worm (and many other animals) with the ability to send signals more rapidly than chemical diffusion, by using localized ion gradients to generate an electrical current down the length of their exterior. There are 302 neurons in the adult hermaphroditic C. elegans, which may seem like a tiny number compared to the hundred billion neurons that humans have in the brain alone, but in fact we have not yet fully deciphered their behavior; how the temperature-remembering behavior mentioned above works is not yet known.
In order to be functional, a neuron must consist of one or more ‘inputs’, a cell body which contains the nucleus and also decides whether or not to take action based on a given pattern of these inputs, and an ‘output’, along which the final signal is sent. If a neuron is capable of receiving input from another neuron, the input portion is a dendrite, so-named because introns in higher organisms often have many, many inputs and these portions of their structure resemble trees. The output portion is called an axon, and is generally non-branching until its terminal region.
Neurons meet at structures called synapses, which are pockets of intercellular fluid into which special signalling molecules called neurotransmitters are released from the axon following a raise in calcium levels, received by surface receptors on the next dendrite, and then recycled. These receptors are often GPCRs or guanylate cyclases, making the evolution of sensory cells like the amphid a straight-forward affair. Reception by GPCRs is enough to start a post-synaptic potential (either excitatory or inhibitory) which will be integrated at the cell body with input from any other dendrites to determine whether or not an action potential is appropriate.
The details of how potentials and synapses work is a favorite subject of physiology and introductory biology courses, and most students are likely already familiar with their inner workings. The links below hopefully contain enough information to subsidize the mending of any discrepancies:
The Nervous System in C. elegans
- WormBook: Synaptic function in C. elegans
- The C. elegans Neural Network
- WormWeb: C. elegans Interactive Neural Network
Neurons (General Reading)
The nervous system is one of the worm’s best-studied subsystems, and may potentially be engineerable to a much greater extent than other systems. “Specification of the nervous system” in WormBook documents a considerable amount of material regarding the lineage of the worm’s neurons, and may help with targeting neurons other than those for which we have already prepared promoters.
One simple way of engineering the worm’s nervous system is through optogenetics, which involves placing light-gated ion channels in the membranes of nerve cells and then stimulating them to induce (or repress) action potentials in a controllable manner. We have endeavored to do this with our initial toolkit; the resultant channels and neuron-targeting promoters can be found on our parts list.
Locomotion: The Consequences of Sensation
Movement of the worm is controlled by interactions between the nervous and muscular systems. About 75 of the worm’s neurons innervate body wall muscles that control movement. The motor neurons of C. elegans are classified into groups by their anatomical position and by the neurotransmitters they release:
| Ventral | Dorsal | |
| Acetylcholine
(excitatory neuron) | VA (backward movement)
VB (forward movement) | DA (backward movement)
DB (backward movement) AS |
| GABA (inhibitory neuron) | VD | DD |
| Other | VC (egg-laying behaviour) |
The neurotransmitters from these motor neurons bind to very specific receptors on the muscle cell’s outer membrane (called the sarcolemma), which accordingly produces localized depolarization or repolarization at the area of binding. When sufficient depolarization occurs, an action potential is triggered, just like in interneurons, and this travels down the sarcolemma.
Muscle Function
Action potentials produced in the sarcolemma eventually travel into the t-tubules, which are structures inside of the muscle cell that exist solely for the purpose of communicating these signals. The t-tubules connect directly to the muscle cell’s endoplasmic reticulum, known as the sarcoplasmic reticulum, where the action potentials alter the conformation of dihydropyridine receptors (DHPR) and Ryanodine receptors (RyR) located thereon. These proteins then work together as voltage-sensitive calcium channels, their conformational change allowing calcium ions to be released from the SR into the muscle fibers.
Muscle fibers consist of a chain of myosin proteins surrounded by six chains of actin proteins. Myosin is a motor protein that changes shape in response to the state of its binding sites and is capable of crawling up an actin filament (in this case, a molecular rope) if conditions are favorable. Six actin filaments are necessary because myosin heads align 60° to each other when stacked. There are two sets of actin filaments anchored to opposite walls of the muscle cell, accompanied by a myosin fiber that spans them, and switches directions in the middle (called the M-line). When the muscle contracts, the myosin chain pulls both sides of the muscle cell membrane towards the middle.
The myosin and actin filaments are not unaccompanied, however. The head domain of myosin is normally blocked from advancing up the actin filaments by tropomyosin, a long molecule that wraps around the myosin filaments and covers their active sites. Tropomyosin is removed by troponin when calcium ions are available; thus, when the sarcoplasmic reticulum releases the necessary calcium ions, muscle contraction occurs. ATP is responsible for providing the energy to rhythmically change the conformation of the myosin heads. Returning to a blocked state causes the myosin fibres to lose their grip, and the actin fibres are pulled away as the cell returns to its natural elastic state.
Locomotive Patterns
The worm’s movement through its environment is a result of systematic and rhythmic contractions and relaxations of the different sets of muscles in its body wall. The predictable, repeated behaviour of this movement is governed by a pattern generator, a neuron responsible for producing the rhythmic signal that causes muscle cells to contract and relax in a predictable fashion. There are probably two pattern generators, one per side, which are out of phase with one another.
The process of studying and mapping when neurons fire and what the consequences of those firings are is called electrophysiology, and it is still largely in its infancy in C. elegans, owing to the tiny size of its neurons. Much more work has been done in the related nematode Ascaris suum, which is larger and has a very similar configuration of its neural net, but there are limitations to this similarity: for example, as of the writing of C. elegans II, it was believed that the cells responsible for pattern generation in Ascaris for regular sinusoidal movement did not correspond to those in C. elegans.
However, what triggers these behaviors is better understood, and to synthetic biologists, the neurons responsible are targets of potentially great interest. The neurons AVB and PVC direct forward locomotion, while AVA and AVD direct backward locomotion. Activating these neurons can induce either direction of movement. This control could be combined with the mutant strain mentioned in this paper as E611 which causes the worm to always keep its head oriented towards a chemical gradient of interest, to allow fixed and predictable radial dispersion from a single source. More complex movement behavior is well-described at length in Gray et al. 2004, especially turns and what triggers them.
G-Protein–Coupled Receptors (GPCRs)

From Wikipedia
GPCRs are common features of eukaryotes and, in multicellular organisms, are one of the major means by which tissues communicate with one another. In C. elegans, they are also used for sensory purposes via the amphid. (We use some of them similarly in our noses.) Many projects have the potential to be highly dependent on GPCR importation and engineered GPCRs, although so far their use at iGEM has not been extensive owing to the high amount of work required to get them to function in prokaryotes.
A GPCR is a transmembrane protein with seven distinctive alpha-helices that serve as transmembrane domains; because of this snake-like structure they are also known as seven-trans-membrane-domain receptors (7TM) or serpentine receptors. The amino terminus faces outward, and in conjunction with the loops of amino acids that extend beyond the membrane, functions as the recognition site. On the inside, the carboxy terminus and the interior loops participate in binding the G protein, a three-subunit structure which is responsible for starting the messaging cascade inside of the cell.
GPCRs are very highly portable between eukaryotes: Teng et al. 2006 showed that some human and mouse GPCRs could be put into C. elegans without modification. Because many organisms have so many GPCRs (humans have about 800 separate GPCR-coding genes; C. elegans has 1200; other related worms may have up to 1600), the evolutionary drift of the interface between these components between species is comparatively small. The primary factor as to whether or not a GPCR will work in a given C. elegans cell is a matter of the availability of the appropriate G protein complex.
The three subunits of G proteins, α, β, and γ, each come in multiple varieties in the nematode, and only certain combinations of these varieties will bind to certain receptors. During signal transduction, the βγ complex is stable and functions as one unit, which simplifies matters slightly, but there are still four possible βγ complexes (two β and two γ subunits), and 21 α subunits. The composition of the βγ complex can be ignored because it does not interact directly with the receptor (only the α subunit does), but projects that involve changing the expression patterns of the α subunits may require bringing gpb-2 or gpc-1 genes with them. (gpb-2 is expressed in only neurons and muscle; gpc-1 is only expressed in certain sensory neurons; gpb-1 and gpc-2 are expressed universally.)
The Alpha Subunits
GSA-1 (analogous to vertebrate Gαs)
- adenylate cyclase (acy-1) —› cAMP —› protein kinase A and cyclic nucleotide-gated ion channels
- vertebrate version can activate calcium channels to contract muscle, inhibit sodium channels in heart
- expressed in neurons and muscles in C. elegans
EGL-30 (vertebrate Gαq/Gα11)
- can couple to vertebrate α1-C adrenergic receptor
- PIP2 —› IP3
- triggers calcium release from internal stores (synapse firing)
- normally localized near axon terminals in acetylcholine-releasing neurons
GOA-1 (vertebrate Gαo)
- counters EGL-30
- expressed in nerves, pharynx, digestive, sex muscles, distal tip cells in hermaphrodites
- partially redundant with GPA-16
GPA-12 (vertebrate Gα12/13)
- predominantly expressed in pharynx, hypodermal cells
- protein kinase C (tpa-1)
GPCRs, G-proteins, and worms
- Weng et al. 2006: Expression of mammalian GPCRs in C. elegans generates novel behavioural responses to human ligands
- WormBook: heterotrimeric G-proteins (where most of this is from)
- WormBook: putitative chemoreceptor families
- GLIDA: search for GPCRs by ligand
- GPCRDB: search for GPCRs by structure, function, or homology
The remaining 17 alpha subunits do not bear significant resemblance to those used in vertebrates, and most likely will not be useful for porting over GPCRs, but neurons with particularly large varieties of alpha subunits such as ASH, ASI, or AWA may still have improved chances. The majority are used heavily or exclusively in sensory neurons, where they may be involved in the decision to enter [[Team:Queens-Canada/reproductive#dauer|the dauer state]. Most of the information below is from WormBase.
GPA-1
- expressed in ADL, ASH, ASI (faint), ASJ, PHA, PHB
- special male expression pattern: also in 1 or 2 pairs of p.c.s. neurons in the tail
GPA-2
- expressed in PVT, PHA, PHB, AIA cells, mu_sph, M1, M5, and I5
- chemotaxis to water-soluable odorants
GPA-3
- expressed in PHA and PHB, and amphid cells, especially AWC and AWA
GPA-4
- expressed in ASI
GPA-5
- expressed in AWA, ASI (faint)
GPA-6
- expressed in AWA, PHB, ASI (faint)
- does not localize to cilia
GPA-7
- expressed in most neurons and all muscle cells
- also neurons in male tail
GPA-8
- expressed in URX, AQR, and PQR
GPA-9
- expressed in ASJ, PHB, PVQ, pharynx muscle, spermatheca
GPA-10
- expressed in ADF, ASI, ASJ, ALN, CAN, LUA, and the spermatheca
GPA-11
- expressed in ADL and ASH
GPA-13
- expressed in ADF, ASH, AWC, PHA, and PHB
GPA-14
- expressed in ASI, ASJ, ASH, ASK, ADE, PHA, PHB ALA, AVA, CAN, DVA, PVQ, RIA, and in vulval muscle
GPA-15
- expressed in ADL, ASH, ASK, PHA, PHB, the distal tip cell, the anchor cell, and many male-specific neurons
GPA-16
- expressed in AVM, PDE, PLM, BDU, PVC, and RIP, and weakly expressed in the pharynx, body-wall muscle, and vulval muscle
GPA-17
- expressed in intestine
- no known use (or consequence for loss)
- similar to Gαq (egl-30)
ODR-3
- expressed in AWA, AWB, AWC, ASH, and ADF
- the AWC neurons consistently express GFP most strongly
- localized to cilia
- the AWB neurons express at lower levels
- the AWA, ASH, and ADF neurons express only weakly
Continue to the Worm's Exterior